
| 23 |
|
Nachdem wir einige Vererbungstatsachen kennengelernt haben, wird es gut sein, uns mit etlichen Vererbungs-Fachausdrücken bekannt zu machen. Im folgenden wird sich der Gebrauch von Fachausdrücken wohl empfehlen, denn man kann sich in der Fachsprache viel kürzer und viel genauer verständigen.
Mancher Leser wird vielleicht sich angeregt fühlen, auch an Fachschriften zur Vererbungslehre sich heranzumachen, und da ist es von Vorteil, wenn man sich die Sprache dieser Vererbungsschriften schon vorher zu eigen gemacht hat.
Die idealste Fachsprache, das sind die Erbformeln, ist uns bereits geläufig.
Das, was bisher reinrassig nannten, also Lebewesen von der Formel AA, aa, RR, rr, nennt , man homozygot, auf deutsch etwa “gleichgepaart„, weil bei solchen Lebewesen der eine Erbfaktor bei der Paarung mit einem gleichen Erbfaktor, zusammen gepaart, kombiniert worden ist, also der Faktor A mit dem Faktor A.
Wenn aber der Faktor A zusammen gepaart wird mit a, dann nennt man das entstehende Individuum heterozygot, auf deutsch “verschieden gepaart„.
Demnach ist homozygot gleichbedeutend mit rasserein (etwa auch mit “Vollblut„) und heterozygot gleichbedeutend mit verbastardiert, gekreuzt im engeren Sinn (wenn man will, “Halbblut„). Man spricht auch von Homozygotie und Heterozygotie.
Für die Erbanlagen oder Erbfaktoren hat man auch das kurze Wort Gen geprägt, das sich wiederfindet im Beiwort genetisch.
Individuen, welche in den Erbformeln übereinstimmen, z. B. alle braunen Schnecken der Abb. 11 (beachte die dabeistehenden Erbformeln) oder die roten Wunderblumen und die weißen Wunderblumen der Abb. 9 nennt man sehr glücklich isogen, auf deutsch “gleich in den Erbanlagen„. Auch die rosafarbigen Wunderblumen der Abb. 9 sind unter sich natürlich isogen, weil sie alle untereinander gleich sind in den Erbanlagen, in der Erbformel Rr.
Die hellen Schnecken sind äußerlich alle gleich, nicht jedoch alle innerlich hinsichtlich der Erbanlagen. Sie sind nicht isogen, und trotzdem sehen wir an ihnen etwas Gemeinschaftliches und möchten für sie einen gemeinschaftlichen Fachausdruck haben. Man nannte sie daher isophän, auf deutsch “gleich in der äußeren Erscheinung„.
Also beim idealen Mirabilis-Typus der Abb. 9 sind alle isogenen Individuen auch isophaen, dagegen nicht beim Schneckentypus, denn hier sind die hellen Schnecken zwar alle isophaen, aber nicht isogen. Die Erbformel des einen Teiles lautet ja AA, die des anderen aber Aa.
Da die Unterscheidung der inneren Veranlagung, sozusagen dem inneren Typus und dem äußerlich verwirklichten Kleid, dem äußeren Typus so überaus wichtig ist, empfiehlt es sich, immer erst anzugeben, welchen Typus man meint. Man hat darum die beiden sehr brauchbaren Fachausdrücke eingeführt Genotypus (Erbanlagen, Erbformeln) und Phänotypus (äußerlich verwirklichtes Kleid).
Wenn beim Schneckentypus in F1 die dunkle Farbe des einen Elters sozusagen ganz unterdrückt erscheint und in der Aufspaltung von F2 die weiße Farbe wiederum vorherrscht, sozusagen dominiert, so nennt man das Dominanz von hell. Von der Eigenschaft Braun sagt man dabei, sie trete zurück oder sie verhalte sich rezessiv.
Die Unterscheidung zwischen dem Mirabilis-Typus und Schneckentypus haben wir oben schon als wichtig betont. Die Vererbungsweise beim Mirabilis-Typus, wo die Bastarde in der Mitte liegen zwischen den beiden Ausgangsrassen, nennt man intermediäre (in der Mitte liegende, Mittelweg-) Vererbung. Eine intermediäre Vererbung liegt auch vor, wenn man rein schwarze Hühner mit rein Weißen kreuzt und in F1 fein schwarzweiß gesprenkelte Hühner auftreten.
Wenn, wie im Schneckentypus, F1 entweder der Vater- oder der Mutterrasse nachschlägt, dann kann man dies alternative Vererbung nennen (auf deutsch etwa die “Entweder-Oder-Vererbung„). Das Wort “alternative Vererbung„ wurde aber auch bisweilen in etwas anderem Sinne gebraucht.
Auch der Praktiker gewöhne sich an die Fachausdrücke: Phänotyp—Genotyp; homozygot—heterozygot; dominant—rezessiv. Aber bitte kein Mißbrauch !
| 24 |
 |
Ausgerüstet mit unserem neuen wissenschaftlichen Rüstzeug gehen wir nun mutig an den nächst schwierigeren Mendelfall. Das etwas größere Maß von Aufmerksamkeit, das dabei aufzuwenden ist, lohnt sich wohl, denn erst in diesen etwas verwickelteren Fällen zeigt sich, wie groß die Dienste waren, die MENDEL der Vererbungswissenschaft und der Züchtungspraxis erwiesen hat. Vor allem auch der Bienenzüchter glaube ja nicht, ohne die Kenntnis wenigstens einiger verwickelterer Fälle sichere Hand in seinen Züchtermaßnahmen zu haben. Überhaupt darf man den Mendelismus nur einigermaßen gründlich anfassen, anderenfalls läßt man am besten die Hände davon. Arbeiten, die “das Mendel’sche Gesetz„, wie ein Bienenzüchtungs-Theoretiker buchstäblich schreibt, nur “ganz nebenbei erwähnen„, gibt es genug.
Abb. 12, welche das Ergebnis von Versuchen W. CASTLE’s an Meerschweinchen darstellt, sei zuerst erläutert. Es handelt sich hier um eine P-Kreuzung von Tieren, die zwei Grundunterschiede (beide allerdings sich auf die äußere Erscheinung, nämlich die Behaarung, beziehend) aufweisen: Schwarz und Hell, Borstig und Glatt. Das F1-Tier muß innerlich Bastard sein, äußerlich erscheint es “rein„ Schwarz und “rein„ Borstig. Die Eigenschaft S Schwarz dominiert also über Weiß (Nichtschwarz) s, und die Eigenschaft Borstig B dominiert uber Glatt (Nichtborstig) b. Wir haben also nicht eine Mittelweg- (intermediäre) Vererbung, sondern ganz die Entweder-Oder- (alternative) Vererbung. Bei beiden Eigenschaftspaaren herrscht Dominanz. Bei dem Versuche sind, nachdem man ein F1-Mannchen mit einem F1-Weibchen paarte (unter Umständen wiederholt), vier äußerliche Haartypen (Phänotypen) aufgetreten. Auf eine uniforme F1-Generation ist also wiederum eine aufspaltende F2-Generation erfolgt. Die Spaltung weist Verwickeltere auf, nämlich:
| 9 | : | 3 | : | 3 | : | 1 |
| Schwarz | Schwarz | Weiß | Weiß | |||
| Borstig | Glatt | Borstig | Glatt |
Man muß also allermindestens 16 Nachkommen züchten, um die Aussicht zu haben, alle Typen auftreten zu sehen. Darum sind auf der Abb. 12 bei der Zeile F2 16 Tiere aufgeführt, die Nummern 1–16.
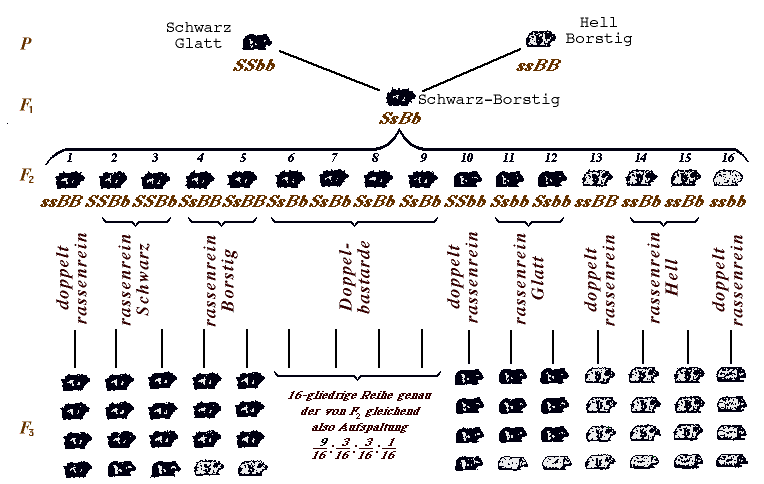 |
Abb. 15. Kreuzung zweier Meerschweinchen-Rassen. Beispiel des Mendelns zweier Merkmalspaare mit Dominanz bei beiden Paaren. [Vergrößerung (19 ko)] |
Erzüchtet man durch Inzucht die F2-Generation, dann sieht man, daß von jedem Typus der F2-Tiere je eines ganz reine Nachzucht hat, es sind dies die F2-Tiere Nr. 1, 10, 13 und 16 (“doppelt rassereine„, d. h. rasserein in bezug auf jedes der beiden Außenmerkmale). Das F2-Tier Nr. 10 z. B. vom Phänotypus Schwarz-Glatt hat eine Nachkommenschaft (die darunter stehende Reihe in der Abb. 12), die alle ohne jedes Aufspalten wiederum Schwarz und Glatt sind.
Das andere Extrem findet sich ebenfalls: von dem häufigsten Phänotypus Schwarz-Borstig finden sich 4 doppeltbastardierte F2-Tiere, nämlich die Nr. 6, 7, 8 und 9, die wiederum eine ganz verwickelte Aufspaltung haben, nämlich genau wie oben 9 Schwarz-Borstig zu 3 Schwarz-Glatt zu 3 Weiß-Borstig zu 1 Weiß-Glatt: Unter diesen Tieren hätte man in der Abb. 12 4 lange, 16gliedrige Reihen zeichnen müssen. Es wurde dies unterlassen, um Raum zu sparen. Die Reihen hätten ja jeweils der 16gliedrigen F2-Reihe in allem geglichen.
Es bleiben dann noch 4 Paare von F2-Tieren übrig, nämlich die Nr. 2, 3; 4, 5; 11, 12; 14, 15. Deren F3-Nachkommen spalten zwar auf, aber jeweils in einem einfachen, vom Schneckenbeispiel uns geläufigen Zahlenverhältnis 3:1. Wenn der Leser an der Hand dieser Beschreibung die Abb. 12 gemustert hat, dann tut er gut, sich in dieselbe noch einmal ohne Text zu vertiefen. Sie ist tatsächlich einfacher, als es auf Grund der Beschreibung den Anschein hat.
Liegt eine Aufspaltung nach Vierteln (= 1/2 × 1/2) vor, dann handelt es sich um eine Einfachbastardierung; nach Sechzehnteln (= 1/4 × 1/4) : um eine Doppeltbastardierung; nach Vierundsechzigsteln (= 1/8 × 1/8) : um eine Dreifachbastardierung.