
| 52 |
|
Wenn irgendwo, dann muß bei der Drohne der Phänotypus am ehesten dem Genotypus entsprechen. Oben handelte das Kapitel „Ein warnendes Beispiel für Züchter“ von der Dominanz. Dort wurde der Züchter darauf aufmerksam gemacht, wie manche Bastarde sich hinter dem Kleide der Reinrassigkeit verbergen. Bei der Drohne gibt es keine Dominanz und kein Verbergen hinter dem Kleid der Reinrassigkeit, bei der Drohne gibt es auch keine intermediäre Vererbung. Denn an einer Eigenschaft des Drohnenäußeren arbeitet ganz ungestört nur ein einziger Erbfaktor, dieser also kann nie von dem Konkurrenz-Paarling vergewaltigt werden, wie etwa beim Schneckenbeispiel der Faktor h vom Faktor H. Es braucht auch nie, wie etwa bei den Faktoren H (Rot) und r (weiß) des Mirabilisfalles ein Mittelweg (Rosa), vereinbart zu werden. Der Drohnengenotypus hüllt sich also nie in falsche Farben oder in Mischlingsfarben, weigstens nie in die eigentlichen Mischlingsfarben der Heterozygotie.
Wenn wir die Drohnen also nach ihrem Äußeren gruppieren, dann werden bei den Drohnen die Gruppen am stärksten gegeneinander abgegrenzt sein.
In dem Falle der Polymerie, wo ein und dieselben Eigenschaften, wie die rote Kornfarbe des Weizens (s. Beispiel S. 81) oder die Ohrenlänge der Kaninchen von mehreren Erbfaktorenpaaren abhängen, da kann es auch bei der Drohne vorkommen, daß Übergangsfarben und andere Kompromißeigenschaften auftreten und die Grenzen zwischen den einzelnen Drohnengruppen verwischt sind. Aber das ist offenbar nur im Falle der Polymerie möglich. Dieser Fall der Polymerie läßt sich also bei der Biene viel leichter als anderwärts feststellen, wo er, wie wir sahen, dem Züchter viel Schmerzen zu bereiten vermag.
Die Genotypen der Eier, welche die Königin K in Drohnenzellen legt, können bis zu fünf Jahren unverändert erhalten bleiben, während in der gleichen Zeit die Genotypen, mit denen Descendentinnen der gleichen Königin K die weiblichen Zellen bedenken, inzwischen bis zu zehnmal durcheinandergewürfelt werden konnten. In dieser langen Zeit wird die betreffende Chromosomengarnitur zwar nie durcheinandergewürfelt, erleidet aber in ihrem isolierten Zustande gar mancherlei Dinge. Wenn irgendwo, so scheint es dem Verfasser, Mutationen zu erwarten wären, dann müßte man sie hier an den ungepaarten Genen sehen (vgl. aber auch Kap. 45).
Wir unterschieden oben Gewinnmutationen und Verlustmutationen, und die Verlustmutationen, die öfters schon beobachtet wurden, unterschieden wir in homozygotische und heterozygotische Verlustmutationen.
Die letztere Unterscheidung hätte bei der Drohne überhaupt keinen Sinn. Von der heterozygotischen stellten wir fest, daß sie leichter auftritt, aber schwerer nachzuweisen ist. Bei der Drohne müßten die Mutationen nicht nur leicht auftreten (mindestens so leicht wie heterozygotische bei den übrigen Lebewesen), sondern auch leicht festzustellen sein. Das würde auch von den Gewinnmutationen gelten, falls sie überhaupt vorkommen.
Da für jede Eigenschaft nur ein Gen vorhanden ist (normalerweise) in der Drohne, was müßte geschehen, wenn eine Verlustmutation dieses einen Gens z.B. für Gesamtkörperfarbe oder für Augenfarbe fehlte ? Offenbar eine ganz erhebliche Störung in der Ausfärbung. Bemerkenswert ist immerhin, daß Bienen–Albinos (ungefärbte Tiere), soviel dem Verfasser bekannt, jedenfalls bei der Drohne häufiger vorkommen als bei der Arbeitsbiene. Das gleiche gilt auch von anderen Mängeln in der körperlichen Ausbildung, z.B. der Einäugigkeit.
Das Experiment am Gen ist eine der ungelösten Aufgaben (Postulate) der Biologie. Theorie und tatsächliche Befunde weisen auf die Drohne (Hymenopterenmännchen, männliche Hummel usw.) hin als ein in mancher Hinsicht geeignetes Versuchstier.
| 53 |
 |
Weniger um zu kritisieren, als um mit Hilfe des „Gegenbeispiels“ (sozusagen) zu klären, wollen wir noch die Ansichten von Imkerpraktikern besprechen, die das erfreuliche Bedürfnis hatten, etwas tiefer zu schürfen. Pfarrer Gerstung, von dessen Interesse an Vererbungsfragen seine Zeitschrift „Die deutsche Bienenzucht in Theorie und Praxis“ Zeugnis ablegt, glaubte auf der Wanderversammlung in Berlin 1913 (auch auf der Beratung über Bienenzuchtfragen im preuß. Landwirtschaftsministerium März 1919) „den einzig richtigen Weg, den die Wissenschaft uns lehrt, dargestellt zu haben für die Heranzucht einer befriedigenden edlen Bienenrasse, und ihn auch aus dem Wesen der Bienenwissenschaft begründet“ (S. 121) zu haben. Seine wichtigsten Ansichten sind:
Die erste der Gerstungschen Ansichten ist richtig, nur muß die Einschränkung ("im Eizustand") wegfallen. Wie wir in Kap. 12 sahen, besitzen auch die erwachsenen Tiere je mehr als eine geschlechtliche Anlage.
Hinsichtlich des Punktes 2 hat Gerstung geirrt. Die Weismannsche Unterscheidung zwischen der Keimbahn (den Geschlechtszellen), welche durch Generationen hindurch das Erbgut wie in einen durchgehenden Kanal weiterleitet, und den Körperzellen, dem Soma, welches zwar von dem durchgehenden Kanal das Erbgut bezieht, aber dann wie eine Sackgasse blind endet, ist in vieler Hinsicht fruchtbar; diese Unterscheidung ist jedoch ohne Bedeutung hinsichtlich der geschlechtlichen Anlagen. In beiden Leitungen, sowohl dem durchgehenden Kanal der Keimbahn als in den Abzweigungen der jeweiligen Körperzellen, befinden sich beiderlei geschlechtliche Anlagen, also die männliche wie die weibliche.
Daß es so sein muß, zeigt ja schon die Unstimmigkeit, zu der Gerstungs zweiter Satz (von mir numeriert L. A.) führen mußte. Die Söhne der dort erwähnten Königin, welche im eigenen königlichen Soma die weibliche Anlage aufgebraucht hat, hätte ihrer parthenogenetischen Nachkommenschaft nichts weiter mehr zu überlassen als eine männliche Anlage. Diese Nachkommenschaft (Drohnen) verbraucht nach Gerstung die überkommene Anlage im eigenen (männlichen) Soma. Was hätten diese Drohnen noch zu reservieren in ihren Keimzellen ? Offenbar nichts im Gegensatz zu Gerstungs eigenen Sätzen 1 und 4.
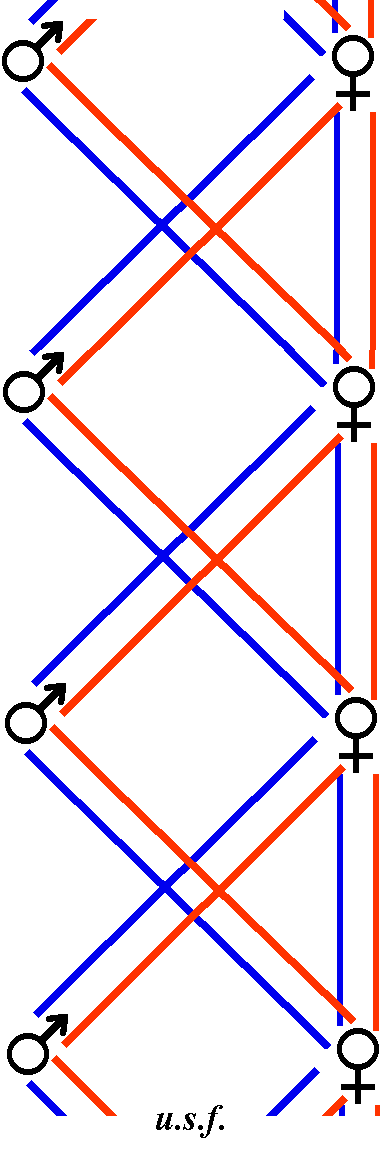 |
| Abb. 20. ——— = Vererbungswege der weiblichen geschlechtlichen Anlagen. ——— = Vererbungswege der männlichen geschlechtlichen Anlagen. |
Ein Schema (Abb. 20) möge den Vererbungsgang der geschlechtlichen Anlagen, so wie er tatsächlich sein muß, vor Augen führen (ein Schema nach Gerstung läßt sich wegen des eben erwähnten Widerspruchs nicht aufstellen).
Aus diesem Schema geht hervor, daß die Drohne tatsächlich beide geschlechtlichen Anlagen empfängt und weitergibt (wie wir oben Kap. 12 angaben). Daß beim weiblichen Geschlecht alles verdoppelt [Wenn oben (Kap. 13) von der einzigen weiblichen „Erbanlage“ die Rede war, so beachte man die Anführungszeichen. Dort ist die Rede davon, daß ein einziger Anlagenkomplex zugrunde liegt, daß aber die jeweilige Lebenslage in einem Falle Arbeitereigenschaften ausprägt, im andern jedoch Königinneneigenschaften.] erscheint (bzw. bei der Drohne dem Weibchen gegenüber alles halbiert), kann uns nint besonders wundern; es ist dies eben eine der merkwürdigen azygoten Vererbungserscheinungen (Kap. 49 ff.). Es würde zu weit führen, wollten wir noch das Verhältnis der sog. sekundären Geschlechtsmerkmale zu den primären im Soma erörtern oder die Frage, wie es kommt, daß zwar Anlagen für beide Geschlechter vorhanden sind in einem Tier, daß aber trotzdem nur ausnahmsweise Zwitter entstehen. Auch die Frage der etwa vorhandene Geschlechtschromosomen bleibt unberührt.
Wenn Gerstung 1919 glaubte, nur anatomische Eigenschaften würden „mendeln“ physiologische aber nicht, so hat er hierin geirrt. Seine früheren Ansichten über die verschiedenen gefärbten Drohnen ein und desselben Stockes wird er heute wohl nicht mehr vertreten [Da die Ansicht aber gewisses historisches Interesse hat (wie schwierig man tat, bevor man die Begriffe Modifikationen und Kombinationen unterscheiden lernte) und eine höchst bezeichnende Beobachtungstatsache (auffallenderes Variieren der azygoten Drohnen) zugrunde liegt, sei sie hier wiedergegeben (Deutsche Bienenzucht in Th. u. Pr. 1913 S. 142f.):
Verschiedenfarbige Drohnen in einem Stock.
Frage: Vorigen Sommer ließ ich mir eine goldgelbe Amerikaner-Königin senden, angeblich rasseecht. Es war ein schönes Exemplar und ging flott in die Eierlage. Die im Herbst und Frühjahr erscheinenden Bienen waren auch schön leuchtend gelb. Im Sommer jedoch wurden dieselben nur mattgelb; es kamen auch immer mehr krainerfarbige Arbeiter zum Vorschein. Die Drohnen jedoch waren verschiedenfarbig: ganzgelbe, nur etwas gelbe und ganz dunkle. Nun heißt es aber, daß eine Bienenkönigin, beliebig befruchtet, immer echtfarbige Drohnen erzeuge. Ein Verfliegen war unbedingt ausgeschlossen. Wie läßt sich dieses Vorkommnis erklären ?
Antwort: Bei der goldgelben Amerikaner-Königin dürfte die Kreuzung zwischen Italiener- Krainer- und Cyprerblut die Hauptrolle spielen. Deshalb ist es auch begreiflichl, daß die männlichen (soll wohl heißen: weiblichen L.A.) Nachkommen derselben leicht in eine der Stammformen zurückschlagen. Um dies zu verstehen, müssen Sie eben die Lehre Mendels studieren (Versuche über Pflanzenhybriden, Leipzig, Verlag von Wilhelm Engelmann). Es würde auch hier zu weit führen, dieselbe, soweit es für die Erklärung Ihrer Beobachtung nötig ist, hier darzustellen. Freilich erklärt der „Mendelismus“ nicht das Erscheinen verschiedenfarbiger Drohnen in einem Stocke, die nachweislich von einer Mutter erzeugt sind. Ähnliche Beobachtungen sind auch früher schon gemacht worden, und man hat stets vor einem Rätsel gestanden, und zwar darum, weil die Drohnen nicht aus einer Kreuzung verschiedenfarbiger Rassen hervorgehen, sondern stets nur Nachkommen einer unbefruchteten Königin oder aus unbefruchtet gebliebenen Eiern sind. Gewisse Leute führen ja diese Erscheinung der Farbenerscheinung zum Beweis dafür an, daß auch die Drohneneier, falls sie von einer befruchteten Königin herstammen, befruchtet sein sollen, was bekanntlich alle wissenschaftlichen Bienenforscher mit immer größerer Entschiedenheit auf Grund der Untersuchung der Bieneneier bestreiten. Bei dieser eigentümlichen Sachlage bleibt nur eine Erklärungsmöglichkeit übrig, welche aber viel Wahrscheinlichkeit für sich hat: Die Färbung der Drohnen ist weniger eine Folge von Vererbung als vielmehr von direkten Einflüssen der Ernährung, des Klimas, der Sonnenenergie usw., denen sie bei ihrer Entwicklung ausgesetzt gewesen ist. Wir hätten es mit einer ähnlichen Erscheinung zu tun wie bei denen, die man unter der Bezeichnung Mimikry in der Naturwissenschaft zusammenfaßt. Wahrend man früher hier und da annahm, daß diese Anpassungsvorgänge absichtlich als Schutzmittel gegen Verfolgung von gewissen Tieren durchgeführt werden, ist man heute durch Versuche mit einem Höhlenmolch zu der Erkenntnis vorgedrungen, daß schon die veränderten Licht- und Farbeneinflüsse, denen man das werdende Tier oder Pflanze aussetzt, imstande sind, Veränderungen in der Oberhautfärbung hervorzurufen, so daß man die Epidermiszellen mancher Lebewesen mit der lichtempfindlichen photographischen Platte vergleichen kann. Bisher ist von diesem Standpunkte aus die Farbenveränderlichkeit der Drohnen noch nicht untersucht worden. Da aber eine andere befriedigende Erklärung nicht vorliegt, so dürfte es schon der Mühe wert sein, einmal in dieser Richtung das forschende Auge und Interesse einzustellen.“].
Gerstung gab 1913 die aus seinen Ansichten folgende Zuchtanweisung : die Zuchtziele der praktischen Imkerei beziehen sich auf weibliche Eigenschaften (s. auch unser Kap. 6). Es vererbt aber „die Drohne die weiblichen Merkmale (weiblichen Eigenschaften als Zuchtziele L.A.) ihrer Mutter, die von ihrem Großvater herstammen“ (S. 120). Also: „Wähle den Zuchtstoff für deine Königinnen nicht sowohl aus den Völkern, deren Eigenschaften dich befriedigen, sondern greife zurück auf das Volk, aus dem die Drohne herstammt, welches die Königin dieses Volkes befruchtet hat“ (S. 120). Oben (Kap. 11) mußten wir einschärfen: „Man unterschätze den Zuchtwert der Drohne nicht“ (S. 23). Hier müssen wir einprägen:
Man über schätze den Zuchtwert der Drohne nicht.
| 54 |
 |
Der rührige Königinzüchter Müsebeck kommt in einem Artikel „Selektion“ in der Leipziger Bienenzeitung 1919 S. 94 zum Ergebnis: Weil man bei verschiedenen Lebewesen, z.B. bei Bohnen, von denen man im Verlauf der Generationen immer die größten „Bohnen“ zur Aussaat benutzte, trotz dieser Auswahl ("Selektion") nach dem Zuchtziele hin, doch dem Zuchtziele nicht merklich näher kam, darum habe man in der Wissenschaft die Selektionstheorie fallen lassen. In Übereinstimmung damit habe auch „die Nachzucht von Hühnern fruchtbarer Legerassen, nach obigen Gesichtspunkten ausgewählt durch neun Generationen,“ keine Steigerung des Eierertrages gezeitigt, und dementsprechend sei „die Herauszüchtung langrüsseliger Bienen nicht möglich“.
Es sei zunächst noch einmal hingewiesen auf die großen Zugeständnisse, die in den Kap. 41 und 44 (Mutationen und Akklimatisation) an gewisse Vererbungstheorien gemacht worden sind, im übrigen sei aber um so nachdrücklicher hingewiesen auf zwei grundverschieden gelagerte Fälle (zwei Fälle, die Müsebeck nicht unterscheidet): den Fall bei erblich einheitlichem (Kap. 54) und den bei uneinheitlichem Material (Kap. 55).
Die Selektion bei erblich (genetisch, genotypisch, vgl. Kap. 23) einheitlichem Material sei erläutert an einem etwas ferner liegenden, aber um so lehrreicheren Beispiel, dem (einzelligen) Urtierchen Paramaecium (Pantoffeltierchen). Die Fortpflanzung geschieht hier gewöhnlich durch einfache Zweiteilung, also durch die gewöhnlichste Zellteilung (nicht etwa Reduktionsteilung !), die das Erbgut nicht etwa umwürfelt, sondern unberührt läßt und gleichmäßig verteilt. Die Schar der Nachkommen ist schon nach wenigen Teilungsschritten sehr zahlreich. Trotzdem die Nachkommen nun isogen (übereinstimmend in der Erbanlage s. S. 45) sind, erscheinen sie gewöhnlich keineswegs isophän (gleich im äußeren s. S. 45), denn ihre Lebenslage ist, wie meist, nicht gleich gewesen.
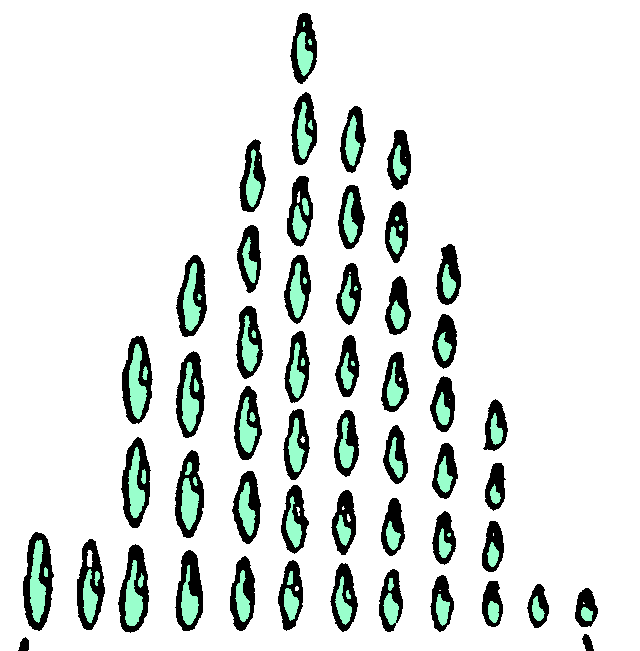 |
||
|
|
|
|
|
|
| Abb. 21. Schematischer Selektionsversuch an erblich reinem Material. Nach BAUR. Jede „dreieckige Gruppe" stellt ein Uhrschälchen Kultur von Paramaecium dar. Die Basis jedes „Dreiecks" versinnbildlicht den „vererbten Spielraum“ (Größentypen). Die gleich großen Kultur–„Geschwister“ sind übereinander angeordnet. Die Durchschnittsgröße ist naturgemäß stets am häufigsten (8 mal) vertreten. Weitere Erklärung im Text. Linke Hälfte: Selektion auf Groß; rechte Hälfte: Selektion auf Klein. |
||
Kann man bei solchen Paramaeciumscharen nun eine Eigenschaft durch Selektion steigern, so daß man mit der Zeit beispielsweise immer größere Paramaecien erhält ? Wir wollen den Versuch nachmachen, der in Abb. 21 dargestellt ist. Wir sondern ein mittelgroßes Paramaecium ab in einem Uhrschälchen mit Nährflüssigkeit und warten, bis durch fortgesetzte Zweiteilung eine ordentliche Schar von (dem Erbgute nach völlig gleicher, in Wirklichkeit aber infolge der Lebenslage verschieden großer) Paramaecien im gleichen Uhrschälchen vorhanden ist (dreieckige Gruppe oben in der Abb. 21). Die mittelgroßen Tierchen sind am zahlreichsten (achtmal) vertreten und in der Mittellinie der Gruppe aufgezeichnet. Nun wollen wir Selektion nach „Groß“ (linke Seite der Abb. 21) und andererseits nach „Klein“ (rechte Seite der Abb. 21) treiben. Wir nehmen ein „selten“ großes Individuum aus dem ersten Uhrschälchen und übertragen es in ein neues Uhrschälchen (Mittel-Zeile links). Von den Nachkommen waren die einen gut gediehen, die anderen kümmerlich. Unser Übertragenes Riesen-Paramaecium hatte nicht seinen Riesenwuchs vererbt, sondern den Spielraum von Groß bis ziemlich Klein (vgl. das Primelbeispiel Kap. 7). Der „Spielraum“ im neuen Uhrgläschen ist genau gleich wie der im alten, ebenso die Durchschnittsgröße. Unverzagt wiederholen wir; ein „einzig“ großes Riesen-Paramaecium wird in ein drittes Uhrschälchen gebracht, und es vermehrt sich dort zu einer neuen Schar (Abb. 21 unten links). Aber am „Spielraum“ und an der Durchschnittsgröße hat sich nichts geändert. Das Zuchtziel: „größere Leibesfülle", haben wir trotz der Selektion nach Groß nicht erreicht. Selektion nach Klein (rechte Seite der Abb. 21) führt ebenfalls nicht zur Verringerung der Körpergröße. Denn nehmen wir vom Ausgangs–Uhrschälchen das allerkleinste Individuum und beobachten dessen Nachkommen (Mittel–Zeile rechts), so finden wir: „Spielraum“ und Durchschnittsgröße sind gleichgeblieben. Wir wiederholen die Selektion nach Klein und sehen dasselbe bei der „dreieckigen“ Gruppe unten rechts. Wir lernen daraus nicht nur, daß ein Riese unter Umständen Zwerge erzeugen kann (dann nämlich, wenn des Riesen Erbgut schlechter ist als sein Aussehen und Phänotypus) und ein Kümmerling umgekehrt wahre Prachtexemplare (wenn nämlich beim Kümmerling äußere Zufälligkeiten die Anlagen zu ordentlichen Erbleistungen verdecken), der Kümmerling hat eben glücklicherweise seine erworbene Eigenschaft nicht vererbt.
Insbesondere lernen wir: unter erblich einheitlichem Material ist jede Selektion erfolglos, gleichgültig, ob diese erbliche Einheitlichkeit besteht unter den Nachkommen eines ungeschlechtlich sich vermehrenden Individuums (die Zweiteilungs-Nachkommenschaft eines Paramaeciums, die ganze Nachkommenschaft einer Kartoffelknolle, an die vielen Obstbäume, für welche die Pfropfreiser oder Stecklinge letzten Endes einem einzigen Obstbaum entnommen worden sind) oder unter den geschlechtlichen Nachkommen eines Tierpaares, das in den Erbanlagen genau übereinstimmte, oder unter den geschlechtlichen Nachkommen von sich selbst befruchtenden Zwitterwesen [Infolge der hier normalen Inzucht verschwinden etwa hereingekreuzte Heterozygote alsbald wieder (vgl. Inzuchtvorarbeit S. 63 und Merksatz 20)], z.B. der Bohnen. Nur pflegt man die ungeschlechtliche Nachkommenschaft eines Lebewesens, ein Klon, die geschlechtlich, erblich einheitliche Nachkommenschaft eine reine Linie zu nennen.
 |
| Abb. 22. Klone von Paramaecien. Nach JENNINGs. Ein ehemaliges Gemisch („Population“) von Paramaecien ist entwirrt und die Tiere, die aus demselben Uhrschälchen stammten, zusammengestellt und je nach der Größe geordnet. Da acht Größenspielräume festzustellen sind, handelt es sich um acht verschiedene Elementarrasse erblich verschiedener Klone. Linie x——x bezeichnet den Größendurchschnitt der ganzen Population, + bezeichnet jeweils die Mittelgroße des einzelnen Klons. |
So können wir uns den Fall der Abb. 22 ebensogut für Bohnen als für Paramaecicn zurechtlegen.
Auf der Abb. 22 sind durch den senkrechten Strich x——x Individuen von einer bestimmten (mittleren) Größe verbunden. Angenommen, wir würden, aus einem großen Bohnensack (geerntet aus den verschiedensten Gärten) 7–8 Bohnen von dieser bestimmten Größe auslesen und deren Inzuchtnachkommenschaft mehrere Generationen studieren auf den vererbten Spielraum, dann würden wir im günstigsten Fall finden, daß 7–8 verschiedene, und zwar erblich verschiedene, Spielräume festzustellen sind, daß also jede der 7–8 Ausgangsbohnen erblich verschieden waren und so zu Ausgangsindividuen für 7–8 „reine Linien" wurden.
Angenommen nun, wir hätten mehr oder weniger zufällig nur die Individuen rechts des senkrechten Striches in unserem Bohnensack gehabt (etwa das Ergebnis eines schlechten Erntejahres), und wir hätten in den kommenden Erntejahren nur von größeren Bohnen Nachzucht erstrebt, dann wären wir offenbar in kürzester Zeit angelangt gewesen bei der reinen Linie der obersten Zeile. Das würde nun aussehen, als hätte die Selektion nach Groß den besten Erfolg gehabt, als hätte die in besseren Erntejahren auftretende erworbene Eigenschaft Groß sich vererbt: — Ist keineswegs der Fall, es wurde lediglich nach und nach eine gute reine Linie isoliert, die deswegen nicht gleich gefunden wurde, weil weniger günstige Außenbedingungen das stets vorhandene gute Erbgut verdeckt hatten — die auch beim nächsten Mißjahr wieder recht unvorteilhaft aussehen mag. Ist aber einmal die beste reine Linie erreicht, dann ist jede weitere Selektion aussichtslos, und höchstens ein Zufallstreffer (das Stoßen auf eine günstige Mutation) konnte zu einem Fortschritt führen.
Also bei erblich einheitlichem Material, beim Klon und bei der reinen Linie, ist nach allem, was wir sicher wissen, Selektion unmöglich.
| 55 |
|
„Selektion“ bei erblich uneinheitlichem Material ist der zweite Fall, den wir kurz abmachen können, weil er schon uns beschäftigt hat beim Weizenbeispiel Kap. 37 S. 81 und dem entsprechenden Fall der Kaninchen mit langen Ohren.
Beim oben erwähnten Bohnenbeispiel (oder Paramaecienfall) war das gute Material, die gute reine Linie (der obersten Zeile von Abb. 22) stets schon in mehr oder weniger Exemplaren vertreten gewesen. Der Züchter war nicht Schöpfer, sondern nur Sucher und Sichter („Selektion“ = Auslese, Sichtung). Beim Fall der roten Weizenkörner, der langen Kaninchenohren, wahrscheinlich auch der hellen (oder dunkeln) Panzerfarbe (und bei der Rüssellänge?) der Biene ist der Züchter mehr als ein Sucher und Sichter, er kann dort sein ein Künstler und „Schöpfer“. Die Mosaiksteinchen der Erbanlagen sind da und dort verstreut in den Erbgütern verschiedener erblich mehr mittelmäßiger Zuchttiere. Indem er aber das Würfelspiel der Chromosomen verfolgt, wie es sich spiegelt in dem Aufspalten nach geeigneter Kreuzung, indem er abwechselnd sucht, sichtet und dann wieder kreuzt und kombiniert, erhält und vermehrt er die Mosaiksteinchen nicht nur, sondern fügt sie zu neuen lebenden, unter Umständen nie gesehenen Kunstwerken zusammen, die im gewissen Sinne seine Schöpfungen sind, Schöpfungen um so erfreulicher, weil nicht nur ihr flüchtiges Auftauchen uns ergötzt, sondern ihr dauernder Besitz (reine Vererbung bei Homozygotie) uns nützen kann [In der freien Natur wären diese Idealwesen unerhörte Zufallstreffer; für den Zürnt er sind sie Werke seines planenden Geistes und seiner geduldigen geschickten Hand.]. Allerdings hat auch dieser Fortschritt einmal ein Ende, wenn nämlich nach dem Weizenbeispiel des Kap. 37 das Lebewesen mit der Formel RRNNHH herausgezüchtet ist. Denn die Nachkommenschaft dieser homozygoten Lebewesen ist erblich rein. Da ist Selektionswirkung zwar unmöglich, aber der Züchter kann mit dem Ergebnis zufrieden sein, selbst wenn ihm auch kein Glückszufall günstiger Mutationen beschieden ist. Ein wahrer weiterer Fortschritt wäre erst dann möglich, wenn das Experiment am Gen geglückt ist (Kap. 52), und wenn damit dieser Triumph den Züchter aus einem „Schöpfer“ von Kombinationen zum „Schöpfer“ neuer Arten und Gattungen machen würde.
Wenn wir uns hier noch unterhielten über den Züchter als Sichter und als Schöpfer, dann haben wir, wie der Aufmerksame wohl gemerkt haben wird, nur noch einmal den grundwichtigen Unterschied berührt zwischen erblichen und nicht erblichen Eigenschaften oder den Unterschied zwischen Modifikationen und Kombinationen. Wir haben darum allen Grund, noch einmal daran zu erinnern, was Züchten heißt (Kap. 2) und uns außer dem Kap. 42 nochmal unseren Merksatz 42 vor Augen zu führen:
„Will der Züchter gute Eigenschaften erhalten, so nehme er sich in acht vor den Modifikationen (z.B. Spielraum bei den Paramaecien) und halte sich nur an Kombinationen und Mutationen.
Will er die guten Eigenschaften auf Kosten der schlechten vermehren, halte er sich insbesondere an die Kombinationen. (Kombinationszucht).
Will er einzelne gute Eigenschaften steigern, dann halte er sich wiederum an die Kombinationen (z.B. Fall der roten Weizenkörner, der langen Kaninchenohren, des schwarzen Bienenhinterleibes), übersehe aber nicht günstige Überraschungen durch Mutationen (Ersatz für das Experiment am Gen !).
Um die Modifikationen kümmere er sich nur insofern, als er seinen Zuchttieren immer die beste Lebenslage zu bereiten sucht; schaden kann das nie“ (s.o. S. 96).
Der Merksatz des vorigen Kapitels:
„Bei erblich einheitlichem Material, beim Klon und bei der reinen Linie, ist nach allem, was wir sicher wissen, „Selektion“ unmöglich“,
ist einer der wichtigsten Sätze der allgemeinen Biologie, und wir hätten mit demselben, so ernüchternd er wirkt, unseren theoretischen Teil der Bienenkunde schließen können. Aber es schadet wohl auch nichts, wenn wir noch kühnen Hoffnungen in diesem Schlußkapitel einigen Raum geben:
Zum „Schöpfer“ wird der Züchter durch das erfolgreiche „Experiment am Gen“ oder durch geschickte Ausnutzung von dessen Ersatz, den „Mutationen", aber auch schon durch die einfachste „Kombination“. Ein Künstler ist der wahre Züchter auf alle Fälle.

|
|
on den in Heft 3 des Archivs für Bienenkunde 1919 aufgerollten Bienenfragen seien hier einige wiedergegeben. Der Verfasser (Berlin-Dahlem, Kaiser Wilhelm Institut für Biologie) wäre für Mitteilungen oder Anregungen äußerst dankbar. Auch unscheinbare Angaben könnten unter Umständen der theoretischen wie praktischen Bienenzüchtungskunde, ja der Vererbungsforschung überhaupt von erheblichem Nutzen sein.