
| 16 |
|
Bei der Befruchtung vereinigen sich väterliche und mütterliche Eigenschaften zum schlummernden Bild des neuen Lebewesens. Wie kommen die Anlagen des Vaters und der Mutter in das Kind? Ganz ohne Zweifel durch das Sperma bzw. durch das Ei, und zwar genauer durch den Spermakern und den Eikern oder endlich noch genauer durch deren Chromosomen. Wenn ein „deutsches“ Bienenei von eineni „italienischen“ Sperma befruchtet wird, dann ist das Ergebnis eine Bastardbiene, auf deren Hinterleibpanzer sowohl die italienische Drohne (hell) als die deutsche Mutter (schwarz) erblichen Einfluß haben. Vom Vater kam ein Erbteil, der sich gelb, von der Mutter ein Erbteil, der sich schwarz auszuwirken sucht; so kam als Ergebnis eine Bastardfarbe zwischen gelb und schwarz zustande.
Nach dem Genannten ist unter den 16 Chromosomen, die vom Vater (Sperma) kamen, eines, das die Anlage für die gelbe Hinterleibsfarbe enthält; unter den 16 Chromosomen, die von der Mutter (Ei) kamen, ist auch eines, das die Anlage für die schwarze Hinterleibsfarbe enthält, oder, was dasselbe sagt, im Tochtertiere ist die Hinterleibspanzerfarbe, diese einzige von fast unzählig vielen Eigenschaften, nicht etwa nur von einem einzigen seiner 32 Chromosomen veranlaßt, sondern von 2. Jede einzelne Eigenschaft steht so in nächster Beziehung zu einem Chromosomen paar. All das macht der Vorstellung eigentlich wenig Schwierigkeiten; es klingt sehr einleuchtend, und man ist verhältnismäßig früh auf diese Gedanken gekommen. Weil es sich aber um grundlegende Auschauungen über Vererbung handelt, hat man sich nicht begnügt, sich diese Dinge, wie ausgeführt, in Gedanken zurechtzulegen, man hat die Aufgabe, all das auch naturwissenschaftlich zu beweisen, ernst genommen. Es macht geradezu Freude, zu verfolgen, wie der Beweis gelungen ist, daß nicht nur die Chromosomenzahl erhalten bleibt, sondern auch die einzelnen Chromosomen-Individuen, also beispielsweise die Rückenfarben-Chromosomen, als solche erhalten bleiben, daß die Chromosomen Paare bilden, und daß der eine der Paarlinge (die Paarlinge sind als solche äußerlich durch die Größe oft kenntlich) dann vom Vater und der andere von der Mutter stammt, daß endlich diese Paarlinge bei der Reduktionsteilung neuerlich getrennt werden, so daß unter den 16 übriggebliebenen Chromosomen alle 16 Paarlinge wenigstens noch vertreten sind (wenn unter den 16 ausgeschiedenen Chromosomen bei Ei und Sperma beide Chromosomen eines Paares, z.B. die beiden Chromosomen der Hinterleibsfarbe, ausgestoßen worden wären, dann wüßte, menschlich gesprochen, der Hinterleib des Tochtertieres, wenn seine Ausbildung überhaupt zustande käme, gar nicht, wie er sich färben sollte!).
Wie mit den väterlichen Chromosomen die väterlichen Eigenschaften in das neue Lebewesen übertragen werden, so mit den mütterlichen Chromosomen die mütterlichen Erbanlagen. Jede Eigenschaft des Jungen beruht auf einem Chromosomenpaar (väterlicher und mütterlicher Bestandteil). Die bei der letzten Befruchtung zu Paaren zusammengetretenen Chromosomen werden bei den kommenden Reifungsteilungen wieder getrennt.
Abb. 8: Die paarweise Zuordnung der Chromosomen auf Grund der Größe erkennbar. Nach Wilson.
| 17 |
 |
Wir haben uns nun einige Vorbegriffe gesammelt über Erbanlagen, über die Träger der Erbanlagen und einiges über die Manöver, welche diese Vererbungsträger ausführen. Wir können nun versuchen, ein Vererbungsexperiment zu beschreiben, wie es ungefähr der zwar spät, dann aber rasch berühmt gewordene Erfinder der höchst bedeutsamen Vererbungsgesetze im Garten des Königsklosters zu Brünn in Mähren begonnen hat. Es handelt sich nämlich um den Augustinermönch Gregor Mendel (1822–1884), dem in 8 Jahren ein kühnes, scharfsinniges, in sich geschlossenes Entdeckerwerk, gelungen war. Einige ungünstige Umstände in seinem Leben und die große Bescheidenheit des Mannes, der seiner Zeit so weit vorauseilte, brachten es leider mit sieh, daß seine Entdeckungen [Erste Veröffentlichung 1865.] bis 1900 begraben blieben. Die moderne Naturwissenschaft rettete insofern ihre Ehre, als nicht weniger als drei gelehrte Botaniker (bezeichnend!) unabhängig voneinander im gleichen Jahre diese Vererbungsgesetze wieder entdeckten: Correns, von Tschermak und de Vries [Die Imkerlehrer, welche vorliegenden Vortrag am 1. Mai 1918 hörten, rechneten sich die Anwesenheit des einen dieser drei Entdecker, die des Herrn Geheimrat Correns, zur besonderen Ehre. Dieser hatte zudem in dankenswerter Weise durch sein Entgegenkommen die Tagung gefördert und prächtiges Anschauungsmaterial zur Verfügung gestellt.].
Die Bienenzucht und die ganze Zoologie muß es sehr bedauern, daß die Vererbungsversuche Gregor Mendels an Bienen nicht nur nicht fortgesetzt wurden, sondern auch alle Aufzeichnungen darüber, die sicher vorhanden waren, der Vernichtung anheimfielen. Mendel war eifriger Imker (Vizepräsident des Vereins Brünn) und ließ es bei seinen Versuchen an Großzügigkeit nicht fehlen. Er hielt 50 Beobachtungsvölker und bezog alle Rassen, die er nur erhalten konnte, aus Europa, Ägypten und Amerika. Zahlreiche Kreuzungen waren ihm gelungen, und wiederholt zeigte er sie in den Sitzungen des Vereins (nach Bateson). Die Schwierigkeiten der Vererbungsversuche bei Bienen bekam er allerdings auch zu spüren. Er suchte bestimmte Kreuzungen mit bestimmten Königinnen zu erzielen mittels abgeschlossener Gazezwinger, hatte aber manchen Mißerfolg. Als er zum Abte seines Klosters gewählt worden war, konnte er nur noch wenig Zeit für Vererbungsexperimente erübrigen. Wir begreifen aber auch, warum er über die Versuche an Bienen nichts veröffentlicht hat. Die Gründe dafür sind dieselben, die uns veranlassen, die Biene nicht als Schulfall der mendelschen Vererbungsweise zu besprechen, sondern erst ein gutes Stück weiter unten.
Zur Darstellung von Mendels Entdeckung sei darum auch hier ein pflanzliches Beispiel gewählt, und zwar das Hauptzuchtobjekt von Correns die Wunderblume, Mirabilis jalapa.
Mendel sagte sich: die Vererbung zu studieren ist weniger interessant da, wo die Eigenschaften stets gleichbleibend weitervererbt werden; lohnender ist es dort, wo Wechsel ist; aber es ist unklug, in den vollen Wirbel des Wechsels, in die ganze bunte Mannigfaltigkeit der Lebewelt sich zu stürzen; besser ist es, auf einen einzelnen Punkt, auf eine einzige, leicht feststellbare Eigenschaft sich zu beschränken. Achten wir z.B., wie die eine Eigenschaft der roten Blütenfarbe mit einer gut davon unterscheidbaren, etwa einer weißen, wechselt. Da liegt es nahe, eine echt rotblühende Pflanze mit einer echt weißblühenden zu kreuzen. „Echt rot“ blühend ist eine Pflanze dann, wenn all ihre Vorfahren schon rein rot waren.
Wenn der Imker in seinem Bienengarten solch reine Pflanzen besitzt, dann kann er ohne weiteres folgenden Versuch nachprüfen.
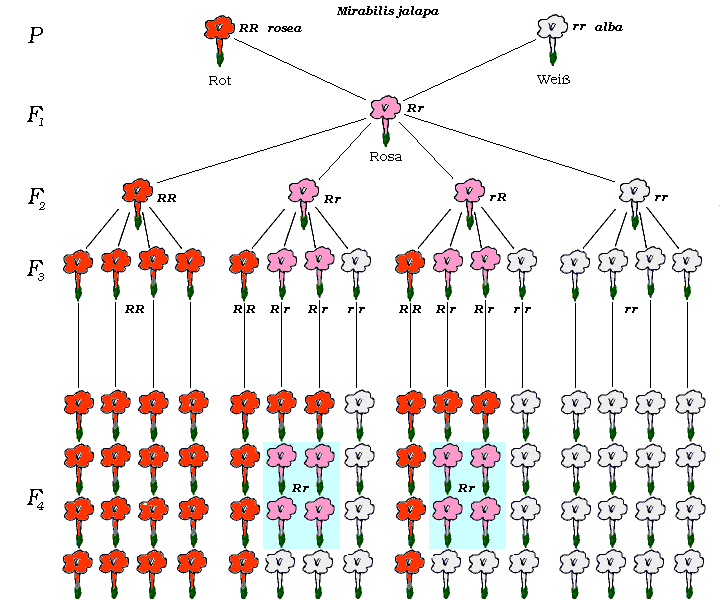 |
Abb. 9. Kreuzung von Mirabilis jalapa rosea und alba. Beispiel des Mendelns von einem Merkmalspaar bei intermediärer Vererbung [Vergrößerung (37 ko)]. |
Nimmt man Blütenstaub von einer rein rotblühenden Wunderblume und bestäubt damit die Narbe [Die Staubgefäße dieser auserwählten Narbenblüte müssen frühzeitig entfernt worden sein, damit deren Blütenstaub nicht den Versuch stört; auch gegen alle Insekten muß die Blüte alsbald abgesperrt werden.] einer rein weißblühenden Wunderblume und sät den aus den betreffenden Fruchtknoten geernteten Samen aus, dann hat man eine Kreuzung rot–weiß, ausgeführt, und man wird finden, als Ergebnis der Kreuzung treten Wunderblumen auf, die sämtlich gleich sind: keine schlägt dem Vater nach und keine der Mutter, alle blühen rosa, also alle liegen in der Mitte zwischen Vater und Mutter. Mit einem einmaligen Vererbungsversuch wird man sich nicht begnügen. Wie wird man jetzt kreuzen ? Man könnte eine Tochterpflanze, eine Rosa–Pflanze, mit einer roten (entsprechend der Vaterpflanze) oder mit einer weißen (entsprechend der Mutterpflanze) oder endlich wiederum mit einer Rosa–Pflanze, also mit einer Schwesterpflanze, kreuzen. Letzteres legt sich nahe. Man nimmt also Blütenstaub eines der erhaltenen Rosa–Bastarde und bringt ihn auf die Narbe einer anderen selbstgezüchteten Rosa–Pflanze. Man kreuzt zwar zwei Bastarde, aber man weiß wenigstens den Stammbaum der beiden. Im nächsten Jahre staunt der Züchter; er bekommt ein buntes Blumenbeet. Ein guter Teil ist zwar wieder rosa, aber rosa ist nicht ausschließlich vertreten (wie man schließlich erwarten könnte; Vater und Mutter waren ja tatsächlich beide rosa), sondern es kommen jetzt auch wieder die Eigenschaften der Großeltern zum Vorschein. Und zwar ergibt eine Zählung: 1/4 sämtlicher Pflanzen des Beetes blüht rot, 1/4 blüht weiß, die Hälfte der Pflanzen, also 2/4 blüht rosa.
Die Elterngeneration, auch Parental–Generation, fachmännisch kurz P-Generation genannt, ergibt eine gleichfarbig (uniform) aussehende Tochtergeneration (erste Filial-Generation, kurz F1-Generation genannt). Kreuzt man diese unter sich, dann ist die Enkelgeneration (zweite Filial-Generation, F2-Generation genannt) nicht mehr einheitlich, sondern sie spaltet auf, wie man sagt, in 1/4 rein väterlich, 1/4 rein mütterlich und 2/4 wie F1.
Voraussetzung des Ganzen ist (wie gesagt)
Hauptergebnis ist
Das sind die grundlegenden mendelschen Vererbungsregeln. Bildlich läßt sich der Versuch leicht darstellen und leicht auch dem Gedächtnis einprägen. In der Abbildung (Abb. 9.) und in allen entsprechenden folgenden stehen in der ersten Zeile die Elter–Individuen (= P-Generation), in der zweiten Zeile ein Vertreter der Tochter–Individuen (F1-Generation), in der dritten Zeile die verschiedenen Vertreter der Enkel–Individuen (F2-Generation). Die F3-Generation ist jeweils der Platzersparnis wegen nicht in wagrechten Zeilen, sondern in senkrechten Reihen angeordnet.
Schon jetzt verstehen wir folgende Grundforderungen für exakte Vererbungsversuche (zunächst theoretischer Art!). Die wissenschaftliche Bienenzüchtung bedarf 1. der Bastardierung reiner, in einer einzigen Eigenschaft leicht unterscheidbarer Bienensorten, 2. Inzucht wäre sehr erwünscht.